研究揭示多倍体植物二型花柱调控机制







文章导读
多倍体植物如何避免异型花柱瓦解?中国科学院华南植物园首次揭秘!通过裂果金花染色体水平基因组组装,科研团队发现S位点超基因在多倍化后仍保持完整功能,关键候选基因SchzAUX22调控花柱发育。研究表明,S位点起源远早于多倍化事件(约5000万 vs 360万年前),证明多倍化不必然导致异型花柱崩溃。这一突破性成果首次从全基因组层面破译分子机制,为被子植物花部多态性进化提供新视角,发表于《新植物学家》。
— 内容由好学术AI分析文章内容生成,仅供参考。
花部形态多样性在被子植物进化与物种分化中具有核心作用。异型花柱(二型或三型)具有提高植物传粉精确度、降低雌雄干扰、促进异花传粉等重要生态功能。全基因组复制或多倍化,对被子植物繁育系统和形态演化具有重要影响。然而,以往对异型花柱分子调控机制的研究都局限于二倍体,对于多倍体植物异型花柱的发生和演化还存在诸多未解之谜。
中国科学院华南植物园首次完成了茜草科四倍体二型花柱植物——裂果金花的染色体水平基因组组装与解析,揭示了其控制花柱异长的S位点超基因,在多倍化后仍保持完整功能。
科研团队基于全基因组测序、比较转录组、形态学、繁殖生物学等多种手段,获得了裂果金花高质量的染色体水平基因组,并鉴定到二型花柱相关的S-locus超基因只存在于短柱花型的个体,呈半合子结构,由包含SchzAUX22在内的四个紧密连锁的基因构成。其中,SchzAUX22是重要的生长素响应因子,被鉴定为调控二型花柱发育的关键候选基因。
研究表明,裂果金花的基因组可能起源于约360万年之前的异源多倍化事件。通过计算S-locus基因和其同源基因的同义替换率(Ks),研究团队发现S-locus的起源时间(约5000万年前),远早于裂果金花多倍化的发生时间。这些证据表明,尽管裂果金花存在由杂交导致的异源多倍化,S位点仍能保持完整并发挥功能,多倍化并不必然导致异型花柱的瓦解。
这是首次从全基因组层面破译多倍体植物异型花柱的分子机制。该成果为深入理解被子植物花部多态性及繁育系统的进化提供了新的视角。
相关成果发表在《新植物学家》(New Phytologist)上。
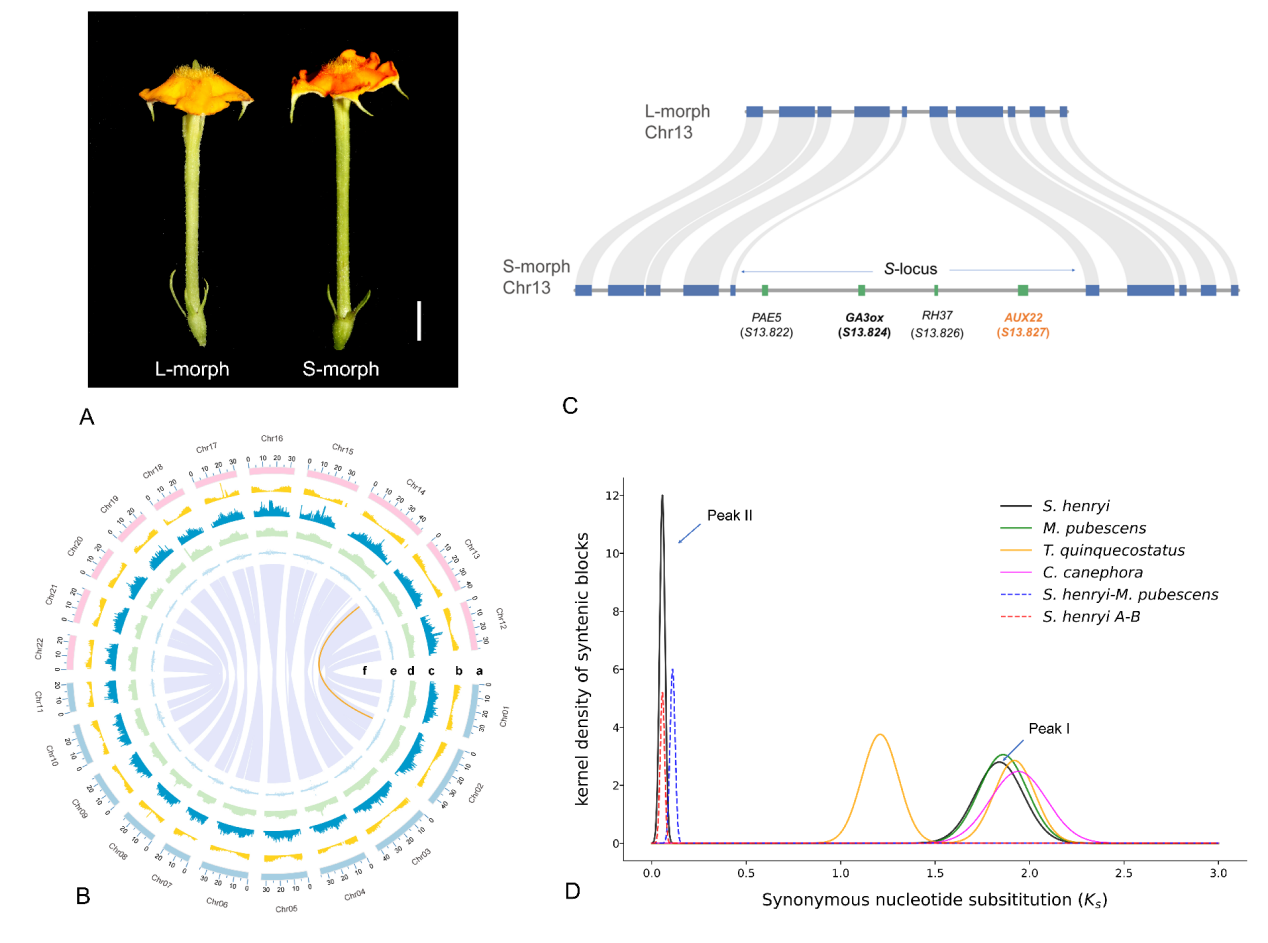
A. 裂果金花长柱型花与短柱型花;B. 短柱型基因组Circos图;C. 半合子位点基因结构示意图;D. 裂果金花与近缘物种旁系同源基因的Ks分布分析。
© 版权声明
本文由分享者转载或发布,内容仅供学习和交流,版权归原文作者所有。如有侵权,请留言联系更正或删除。





相关文章




暂无评论...