哺乳动物精子鞭毛长效运动研究取得进展







文章导读
你可能从未想过,精子能长距离游动靠的不是蛮力,而是精密到分子级的“自我修复”系统。我们一直以为蛋白质折叠只发生在细胞体,但最新研究发现,哺乳动物精子鞭毛内部竟藏着一个活跃的“原位翻译—折叠”流水线。那个在电镜下长期无法识别的桶状结构,竟是特化的TRiC伴侣复合体,它就地捕获新生蛋白,实时折叠,持续替换运动中损耗的部件。更惊人的是,这套机制在斑马鱼等体外受精生物中完全不存在。这意味着,陆生繁殖的代价,是一套专为持久战设计的分子维保系统——而它的崩溃,可能正是许多男性不育的真正根源。如果修复过程出错,身体根本意识不到,但生育能力已在无声中流失。
— 内容由好学术AI分析文章内容生成,仅供参考。
精子鞭毛为含典型“9+2”轴丝的特化动纤毛,其摆动靠精细结构协同驱动,相关骨架蛋白依赖特有大分子伴侣复合体辅助折叠,且该功能异常关联男性不育。从水生体外受精到陆生哺乳动物体内受精的演化,让精子需具备持久游动能力,但其鞭毛长期续航与稳定工作的分子机制有待阐明。
近日,中国科学院分子细胞科学卓越创新中心、生物物理研究所联合大连化学物理研究所等,依托冷冻电镜、原位冷冻电子断层扫描、原位交联质谱和功能实验,揭示哺乳动物精子鞭毛轴丝中连接辐射轴RS1与RS2的桶状结构是精子特异性的TRiC伴侣素复合体,证明精子鞭毛内存在蛋白质原位翻译,且TRiC促进新生蛋白质的折叠并为鞭毛运动性所需,提出哺乳动物精子鞭毛通过蛋白质“原位翻译—原位折叠”进行自我维护来维持持久运动能力的新机制。
此前,冷冻电子断层扫描研究发现,哺乳动物精子鞭毛轴丝比上皮组织动纤毛轴丝在辐射轴RS1与RS2之间多出一个桶状密度(RRB),且该密度在斑马鱼、海胆等体外受精动物的鞭毛中不存在。研究通过非标记定量质谱分析牛和小鼠精子鞭毛发现,分子伴侣素TRiC的8个亚基均富集于鞭毛存在睾丸特异性亚基CCT6B。研究随后解析牛精子鞭毛TRiC的冷冻电镜结构发现,其具有典型的TRiC双环八聚体构象,并包含CCT6A和CCT6B,表明精子鞭毛存在特异性的TRiC复合体。
团队进一步解析小鼠精子鞭毛轴丝96纳米重复单元的原位结构,确认RRB即为TRiC。它通过三个连接网络以特定极性锚定于RS1与RS2之间,并与内、外动力臂及N-DRC等关键运动调控元件有联系。这种多重锚定方式不仅保证TRiC在鞭毛高速摆动中的稳定性,也可能通过影响上述轴丝运动调控组分,参与调控鞭毛运动。原位RRB-TRiC的两个腔室均出现额外密度,其中靠近RS2侧的密度类似经典底物的结合位点,靠近RS1侧的密度则与辅因子Plp2位置相符,说明原位RRB-TRiC并非静息构象,而是处于“底物接收/折叠准备”的活性状态。
研究还在牛和小鼠鞭毛中检测到核糖体、翻译起始因子、氨酰-tRNA合成酶等翻译机器的组分。嘌呤霉素代谢标记实验证实,小鼠鞭毛内存在新生多肽链合成,且断头鞭毛中仍可检测到嘌呤霉素掺入,从而支持鞭毛内存在“原位翻译”的观点。细胞内交联质谱显示,TRiC亚基和核糖体亚基均与多种轴丝结构蛋白发生交联,且存在交集。这些发现揭示了鞭毛中翻译出的新生轴丝蛋白由RRB-TRiC接手折叠的偶联机制。
研究显示,靶向抑制TRiC的ATP酶活性,可抑制小鼠精子鞭毛摆动,但不影响缺乏RRB结构的斑马鱼精子运动,证实TRiC对维持哺乳动物精子运动具有重要作用。基于此,研究发现,在持续机械运动中受损的轴丝组分可通过鞭毛内原位合成、邻近TRiC捕获并折叠的方式及时替换,从而维持鞭毛长期稳定摆动。
该研究赋予了哺乳动物精子鞭毛特有RRB结构明确的分子身份与功能内涵,解释了哺乳动物精子“长续航”运动的结构基础,为理解伴随脊椎动物登陆而出现的、繁殖方式从体外受精到体内受精的转变中精子鞭毛的适应性演化机制提供了新见解,并提示TRiC功能异常或鞭毛内局部翻译与折叠受损或影响精子运动能力并导致男性不育。
相关研究成果在线发表在《分子细胞》(Molecular Cell)上。研究工作得到国家自然科学基金委员会和中国科学院等的支持。
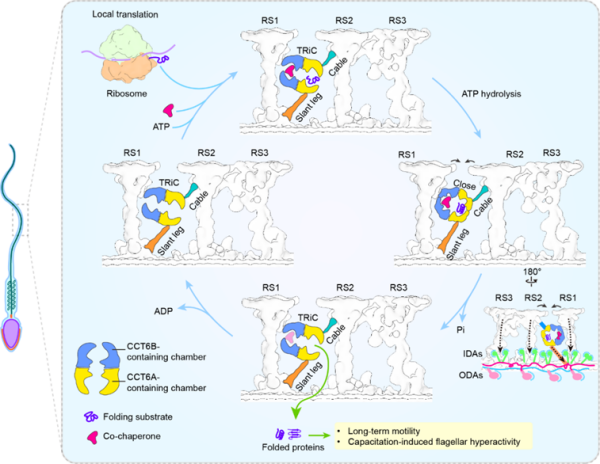
哺乳动物精子鞭毛中“原位翻译—原位折叠蛋白质”的偶联模型
© 版权声明
本文由分享者转载或发布,内容仅供学习和交流,版权归原文作者所有。如有侵权,请留言联系更正或删除。





相关文章



这种基础科研虽然枯燥,但真的解决了大问题😭。
又是看不懂系列,但感觉对人类生育很有用。
所以以后治不育是不是得盯着这个蛋白看?
从水生到陆生演化出这套机制,大自然真神了。
中科院又搞大动作,这波必须给个赞👍。
原来不育可能是这个TRiC复合体在捣乱啊。
看着这些专业名词头都大了,不过感觉挺厉害的。
“原位翻译”是啥意思?有人能给小白解释下吗?
以前只知道备孕难,没想到微观机制这么复杂。
这研究太硬核了,精子居然能自己修零件?